
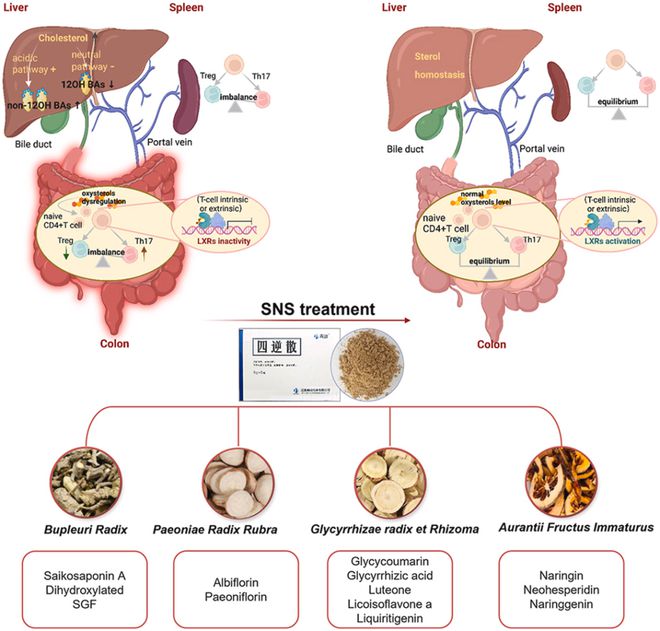
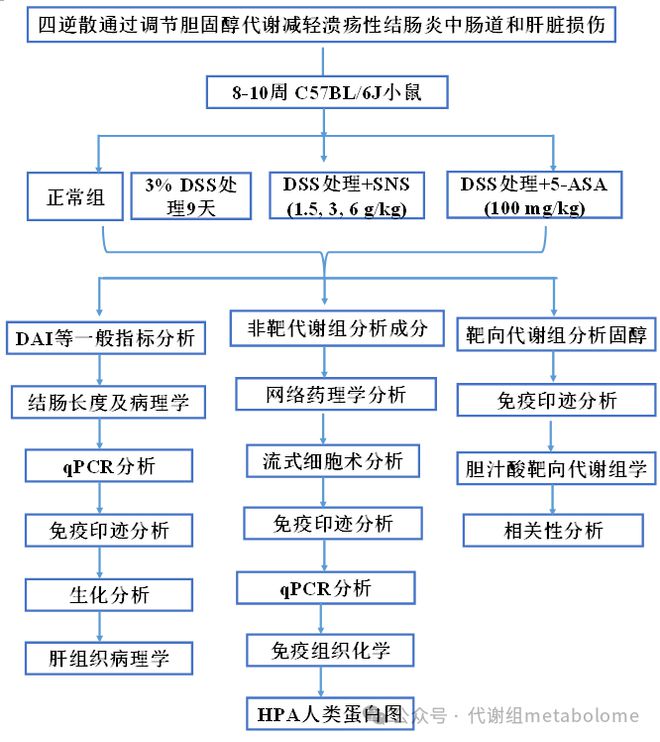
四逆散(SNS)是一种源自《伤寒论》的中药方剂,经过数千年的临床实践,被认为对治疗炎症性肠病有效;然而,其生物活性成分和潜在机制尚不清楚。本研究旨在评估SNS在右旋糖酐硫酸钠(DSS)诱导的小鼠结肠炎模型中改善溃疡性结肠炎(UC)和相关肝损伤的作用,探索其生物活性成分和潜在机制,研究发现SNS(1.5、3、6g/kg)以剂量依赖的方式降低疾病活动指数(DAI)评分,保护肠黏膜屏障,抑制促炎细胞因子产生,改善肝脾肿大,减轻肝损伤。我们在SNS(6g/kg)处理的结肠炎小鼠结肠鉴定出22种成分,按相对含量排名的前10种成分被视为SNS的潜在有效化学成分,并用于网络药理学研究。SNS的疗效是通过减少Th17细胞分化、恢复结肠和脾脏中Th17/Treg细胞稳态来介导的。我们还发现SNS通过上调结肠中CH25H、CYP27A1蛋白的表达来调节25-OHC和27-OHC的浓度,从而影响肝X受体(LXR)的表达和活性,最终影响Th17分化和Th17/Treg平衡。此外,SNS抑制了UC小鼠肝脏胆固醇的增加,逆转了胆汁酸(BA)合成向酸性途径的转变,从而降低了非12-OH BA在总胆汁酸(TBA)中的比例,进一步改善了结肠炎和伴随的肝损伤。本研究基于改善肠道和肝脏损伤的显著作用,为SNS开发成为一种抗结肠炎中药处方奠定了基础,并揭示了胆固醇衍生物,即氧化固醇和胆汁酸,与SNS抗结肠炎作用的机制密切相关。
3.SNS调节结肠“氧固醇LXRs”信号传导,恢复Th17/Treg稳态;
为了研究SNS对急性肠道炎症的影响,健康的C57BL/6J小鼠用3%DSS处理9天,每天口服两次SNS(1.5、3、6g/kg)和5-氨基水杨酸(5-ASA,100mg/kg),持续9天(图1A)。与DSS组相比,SNS(3.6g/kg)以剂量依赖的方式显著降低了以体重减轻、腹泻和出血为特征的DAI评分(图1B-D)。我们通过组织病理学分析进一步研究了SNS对肠黏膜屏障的保护作用。SNS显著缓解了结肠炎小鼠的黏膜溃疡、炎性细胞浸润、隐窝损伤和表面上皮破坏(图1E)。与DSS组相比,SNS(6g/kg)处理的结肠炎小鼠结肠中IL-1β、TNF-α和IL-6等促炎介质的产生受到抑制(图1F-H)。此外,SNS(6g/kg)增强了occludin的表达,occludin是一种代表性的紧密连接蛋白,在DSS组中严重受损(图1I),并减少了结肠氧化应激(补充图S1)。总之,这些结果表明,SNS(3和6 g/kg)对DSS诱导的结肠炎的缓解效果优于5-ASA在100 mg/kg剂量下的缓解效果。
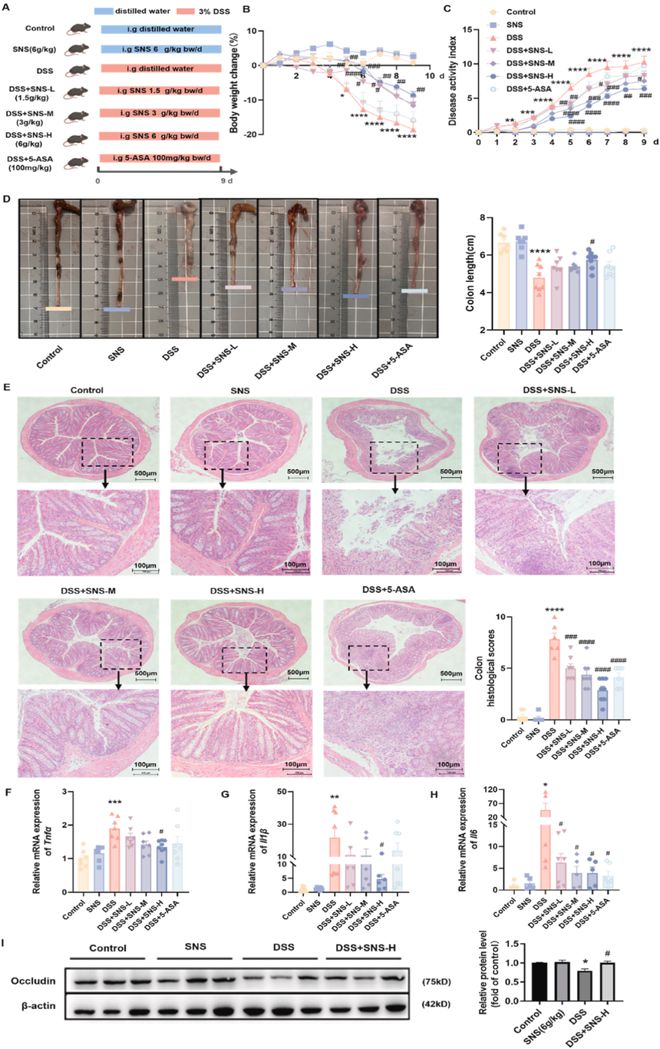
图1 SNS缓解了DSS诱导的结肠炎的症状和结肠炎症。(A)SNS药效学研究示意图。雄性C57BL/6J小鼠被喂食补充有3%DSS的饮用水以诱导结肠炎。分别口服SNS(1.5、3、6g/kg)和5-ASA(100mg/kg),每日两次。(B-C)不同组体重和DAI的每日变化。(D)结肠的长度。(E)HE染色结肠组织的代表性图像(上:40倍放大;下:200倍放大)(n=6)。(F-H)采用RT-qPCR检测炎性细胞因子Il1β、Tnfα、Il6的mRNA表达太阳成集团tyc45668cn。(I)通过蛋白质印迹法检测紧密连接蛋白occludin的表达(n=6)。数据以平均值±SEM表示(除非另有说明,否则n=8-10)。与对照组相比,*P<0.05;**P<0.01;***P<0.001;****P<0.0001。与DSS组相比,#P<0.05;##P<0.01;###P<0.001;####P<0.0001。
由于结肠炎通常伴有其他器官损伤,因此我们还分析了重要器官指标以评估器官损伤。SNS以剂量依赖的方式抑制结肠炎小鼠的肝脾肿大(图2A,B),而其他器官指标在组间没有显示出显著差异(补充图S2)。此外,我们通过体重、结肠长度和重要器官指标评估SNS的体内安全性,SNS处理后我们没有观察到显著变化,这表明SNS在小鼠体内是一种安全的药物。除了丙氨酸转氨酶(ALT)、天冬氨酸转氨酶(AST)活性较高外,DSS组还出现肝脏病理变化,但SNS可以显著降低这些指标(图2C-E)。一般来说,与DSS组相比,SNS可以降低结肠炎小鼠轻度至中度肝损伤的百分比(图2F)。肝脏的病理参数与结肠和DAI中的炎性细胞因子显著相关(图2G),表明SNS对UC的改善可能与其对伴随性肝损伤的调节作用有关。
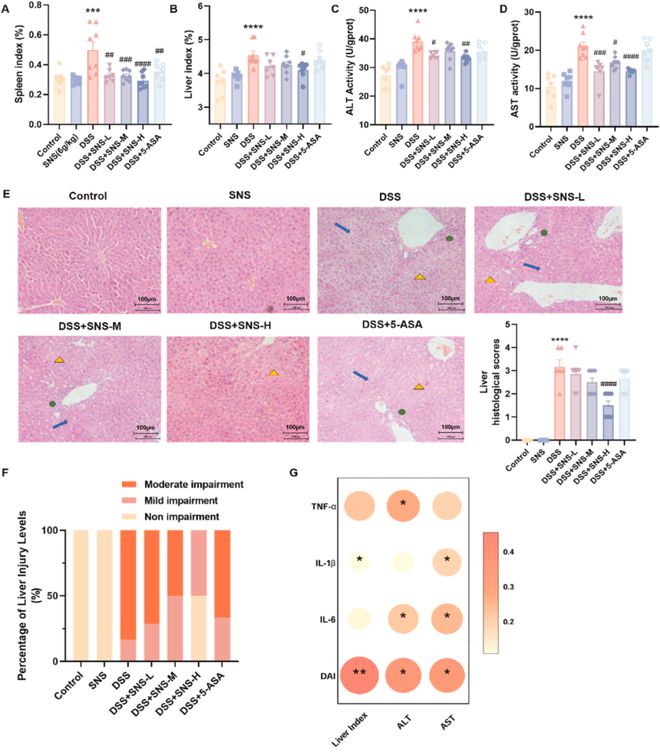
图2 SNS减轻了UC小鼠的肝损伤和脾脏肿大。(A)脾和(B)肝指数;(C)肝匀浆中AST和(D)ALT活性;(E)通过H&E染色(放大200倍)检测组织学变化:绿色圆圈表示炎性细胞浸润,蓝色箭头表示肝索排列紊乱,黄色三角形表示肝细胞凋亡(n=6)。(F)非/轻度肝损伤(0≤2分)、中度肝损伤(2≤4分)、重度肝损伤(4≤6分)的百分比;(G)肝脏和结肠病理参数的Spearman相关性分析。点的大小越大,颜色越深,相关系数越大,标记为*的点表示p0.5,**表示p0.01。数据以平均值±SEM表示(除非另有说明,否则n=8-10)。与对照组相比,*P<0.05;*P<0.05;**P<0.01;***P<0.001;****P<0.0001。与DSS组相比,#P<0.05;##P<0.01;###P<0.001;####P<0.0001。
我们采用HPLC-IT-TOF-MS法对SNS的化学成分进行鉴定和分析。正负离子流图如补充图S3所示。如补充表S7所示,HPLC-IT-TOF-MS以正离子模式或负离子模式对84种植物化学物质进行了表征。然后,我们在SNS(6g/kg)处理的结肠炎小鼠结肠中测定SNS中特征的植物化学物质及其相应的体内代谢产物,成功鉴定出22种化合物(图3D)。相关信息,包括保留时间、加合离子类型和碎片离子,详见补充表S8。这些化合物根据其相对含量进行排名,前10名化合物被视为SNS的潜在有效化学成分(图3E)。
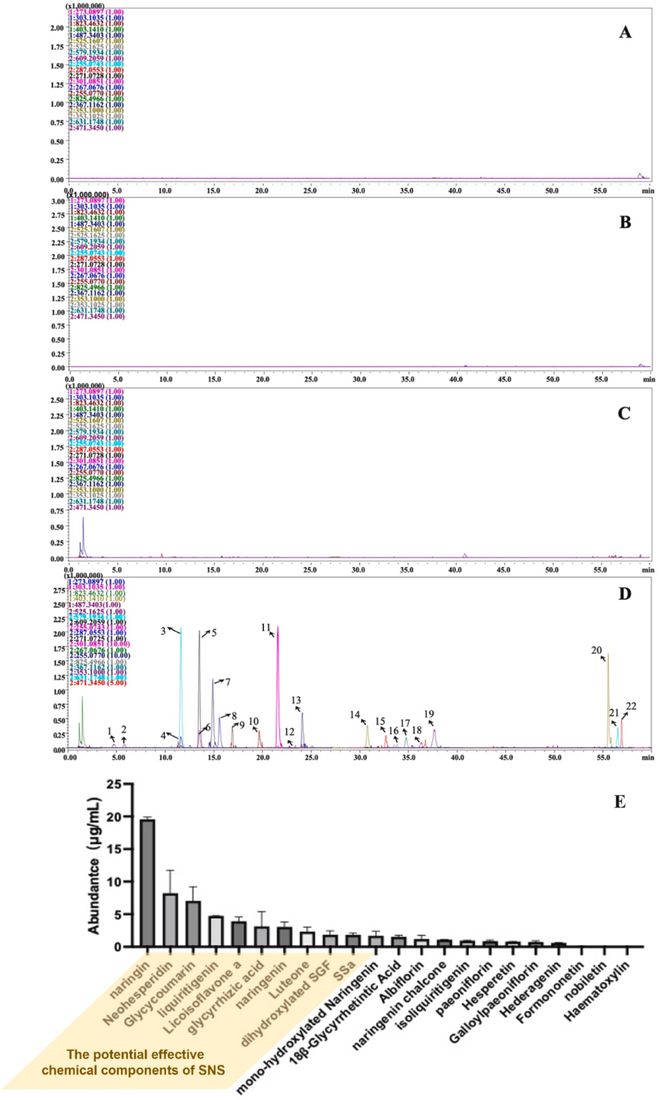
图3 通过LC-IT-TOF-MS鉴定结肠炎小鼠的结肠成分。(A)仪器背景;(B)空白样品;(C)结肠炎组;(D)SNS(6g/kg)给药组;(E)22种来源于SNS的化合物,根据其在结肠中的相对丰度进行排名。
我们利用SNS潜在的有效化学成分构建网络药理学,分别收集了与10种化合物相对应的110个靶点和5759个溃疡性结肠炎靶点,取其交集得到70个交叉靶点,作为SNS改善UC的潜在靶点(图4A)。我们应用Cytoscape软件构建了70多个基因的PPI网络(图4B);基于MCC算法对前5个核心靶点进行了过滤,这些靶点可能是SNS发挥作用的重要靶点(图4C)。根据基于核心靶点的KEGG分析,两条相似的富集通路,即前20条通路中的Th17细胞分化和IL-17信号通路(图4D)引起了我们的注意,初步证据与我们的假设一致,但有待进一步实验证实。
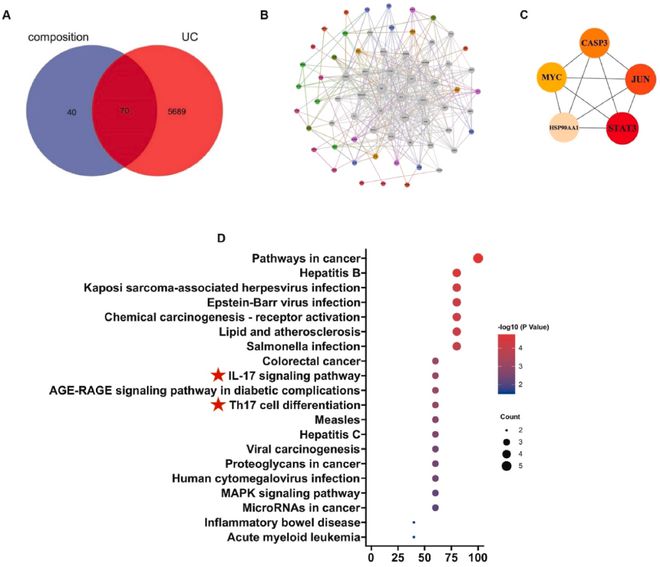
图4 基于SNS关键药效物质的网络药理学。(A)SNS和UC关键药效物质的潜在靶点维恩图;(B)药品-疾病靶点PPI网络建设;(C)PPI网络中基于MCC算法获得的5个核心靶点;(D)基于核心靶点的前20种KEGG信号富集途径。
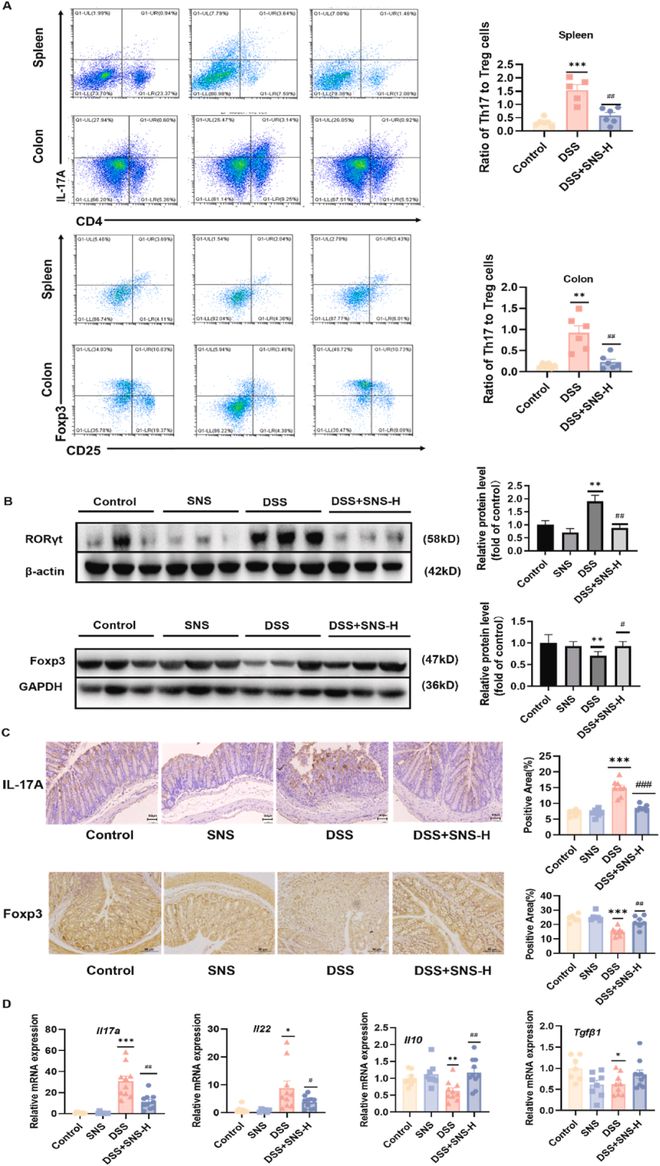
Th17细胞是从CD4+T细胞分化而来的细胞亚群,它们的分化伴随着Treg细胞比例的降低。与DSS组相比,SNS增加了结肠炎小鼠脾脏和结肠固有层中Treg细胞的比例,减少了Th17细胞的比例(图5A)。RORγt和Foxp3分别是Th17和Treg分化的主要转录因子,其蛋白表达代表了SNS对Th17/Treg平衡的类似调节(图5B);这些结果通过针对结肠组织IL-17A和Foxp3染色的免疫组织化学分析得到了进一步证实(图5C)。此外,DSS组Th17细胞的标志性细胞因子如IL-17A和IL-22上调,相应的Treg特异性细胞因子下调(图5D)。总之,充分的证据表明,SNS可以阻断Th17的分化,同时促进结肠炎小鼠的Treg分化。
核心靶点KEGG通路富集分析结果表明,Th17细胞分化可能是SNS改善UC的潜在机制,这得到了实验结果的支持。然而,预测的核心靶点在各种草药和疾病中得到了重现,这引发了人们对这些预测准确性的担忧。人类蛋白图谱(中的免疫细胞表达簇表明,STAT3在免疫细胞中的特异性较低,而LXRs(介导Th17分化的负调控并促进Treg细胞分化的核受体)在T细胞中比STAT3更具特异性(图6A)。接下来,我们测试了SNS对LXRs在体内表达和活性的影响。免疫印迹结果显示,SNS显著上调了DSS抑制的LXRs的蛋白水平(图6B)。LXR活性由Abca1和Npc1l1的mRNA水平指示。ABCA1是LXR的靶基因,其表达随着肝脏X受体(LXR)活性的上调而增加,而NPC1L1的表达则受到LXR激活的抑制。结果表明,SNS给药显著增加了Abca1的表达,抑制了Npc1l1的表达,表明SNS促进了LXR的活性(图6C)。
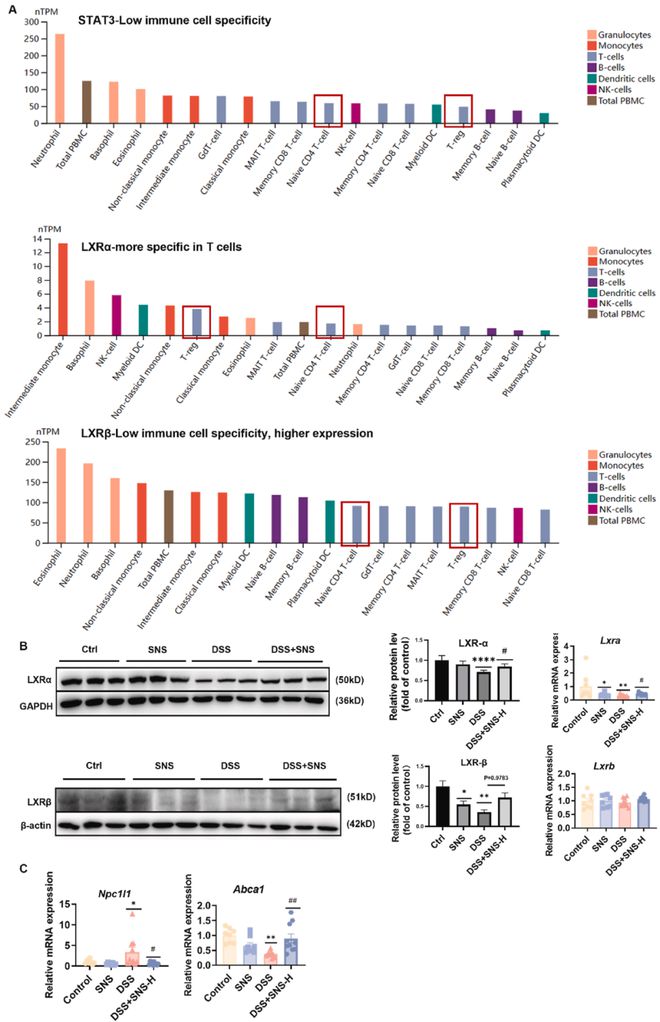
图6 SNS对LXR的表达和存活具有调节作用。(A)STAT3、LXRs表达(RNA)在免疫细胞类型中的分布(图片来源:HPA人类蛋白图谱数据库)。(B)通过蛋白质印迹法检测LXRs的表达(n=6);(C)RT-PCR检测Npc1l1、Abca1的表达(n=8-10)。数据以平均值±SEM表示。与对照组相比,*P<0.05;与对照组相比,**P<0.01;与对照组相比,***P<0.001;与对照组相比,****P<0.0001;与DSS组相比,#P<0.05;与DSS组相比,##P<0.01。
氧化固醇作为LXRs的内源性激动剂,介导幼稚CD4+T细胞分化为Th17/Treg谱系。因此,我们对结肠中的氧化固醇LXR信号传导进行了研究。我们通过LC-MS/MS测定了结肠中作为LXRs内源性配体的氧化固醇的含量。尽管LXRs的拮抗剂4β-OHC在各组之间保持不变,但DSS组中作为LXRs激活剂的27-OHC和25-OHC显著降低(图7A)。与此一致,结肠中氧化固醇合酶、CYP27A1和CH25H的mRNA和蛋白质水平均降低(图7B,C)。DSS组CYP7B1在mRNA和蛋白质水平上的表达均升高(图7D),从而介导了27-OHC和25-OHC的代谢,产生了7α,27-diOHC和7α,25-diOHC,进一步导致27-OHC、25-OHC水平减少。综上所述,上述结果表明,结肠炎小鼠的氧化固醇LXRs信号传导受损,而SNS给药改变了氧化固醇-LXRs信号传导,可能有助于恢复Th17/Treg平衡。
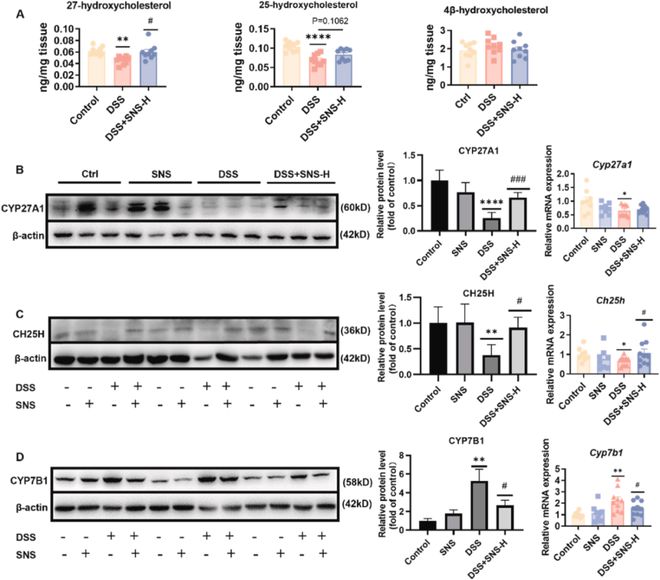
图7 SNS恢复了结肠中LXR的内源性配体水平。(A)以d7-5α、6α-环氧胆甾烷醇和d6-27-羟基胆固醇为内标,采用LC-MS/MS法测定结肠中几种已报道的LXR配体氧化固醇(n=8-9);(B-D)通过蛋白质印迹(n=6)和RT-qPCR(n=8-10)检测了氧化固醇合成的主要代谢酶。数据以平均值±SEM表示。与对照组相比,*P<0.05;**P<0.01;***P<0.001;****P<0.0001。与DSS组相比,#P<0.05;##P<0.01;###P<0.001;####P<0.0001。
我们的实验表明,SNS显著减轻了结肠炎小鼠的肝损伤。我们进一步探讨了SNS抗肝损伤作用的机制。在本研究中,我们在DSS组中观察到肝脏中T-CHO、TBA的异常升高(图8A、B),这暗示了甾醇稳态的破坏。在肝脏中,胆固醇通过中性和酸性合成被催化为BA,据报道,BA代谢异常与肝损伤和包括IBD在内的消化系统疾病有关。我们在DSS组也观察到肝脏BA谱的破坏,而SNS恢复了这一变化。我们发现α-鼠胆酸(α-MCA)、β-鼠胆酸(β-MCA)、牛磺鼠胆酸(T-MCA)、熊去氧胆酸(UDCA)和猪去氧胆酸(HDCA)显著增加,同时非12-OH BA在总BA中的比例增加(图8C,D),表明结肠炎小鼠肝脏从中性途径转变为酸性途径。为了证实BA水平的变化,我们进一步确定了BA代谢途径中关键酶在mRNA和蛋白质水平上的变化。DSS组中性合成途径的关键酶CYP7A1和CYP8B1的表达降低(图8E)。启动酸性途径的StarD1在DSS组中显著增加(图8F),表明结肠炎小鼠肝脏中的酸性代谢途径被激活。非12-OH BA的增加可能有助于UC的进展,因为α-MCA、β-MCA和HDCA与结肠炎性细胞因子和DAI呈正相关(图8G)。结果表明,SNS调节BA合成中的关键酶,逆转UC小鼠BA合成向酸性途径的转变,从而改善肝损伤和结肠炎。
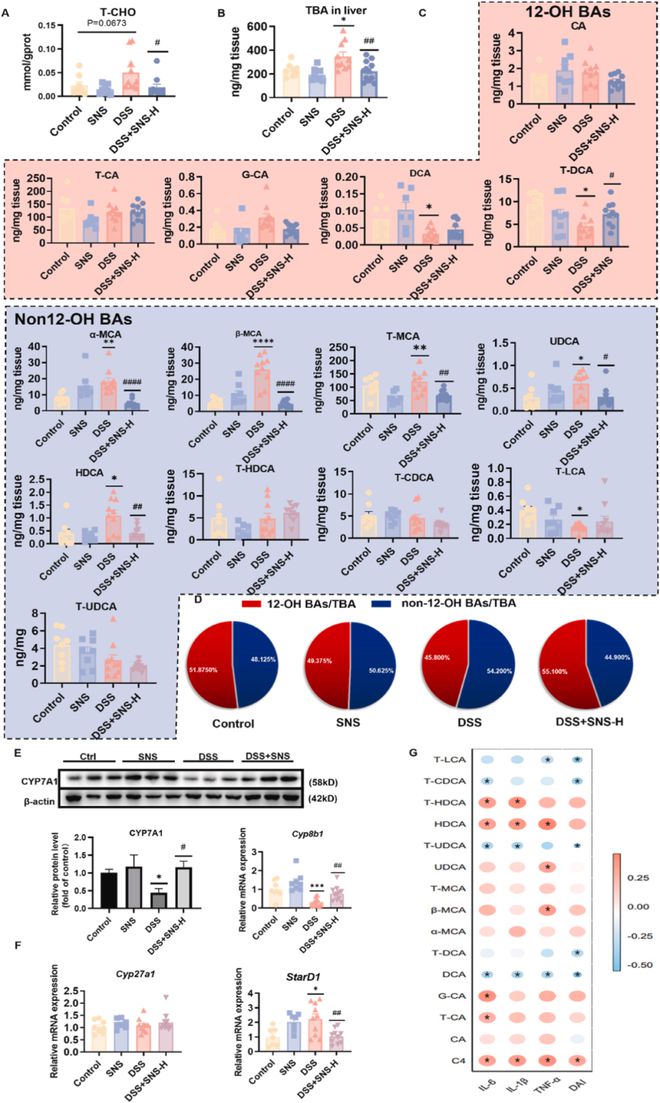
图8 SNS逆转了UC小鼠BA合成向酸性途径的转变。(A)肝脏中的T-CHO;(B)T-BA按单个BA含量的总和计算;(C)以CA-d4、DCA-d4、LCA-d4为内标,采用LC-MS/MS法测定肝脏中BAs水平;(D)肝脏中12-OH BAs的百分比;(E-F)通过蛋白质印迹和RT-qPCR检测BA合成中的关键酶(n=6);(G)热图显示了结肠DAI/炎性细胞因子和肝脏各种BA的Spearman分析。蓝色表示负相关,点越小,颜色越蓝,相关系数越小;红色表示正相关,点越大,颜色越红,相关系数越大,标有*的点表示p0.5。数据以平均值±SEM表示(除非另有说明,否则n=8-10)。与对照组相比, *P<0.05;**P<0.01;***P<0.001;****P<0.0001。与DSS组相比,#P<0.05;##P<0.01;###P<0.001;####P<0.0001。
UC的临床症状包括腹痛、腹泻、便血,有时还会出现肠外并发症。在并发症中,肝胆并发症是最常见的。因此,同时改善肝脏和肠道症状是UC的治疗靶点之一。
SNS是基于“疏肝理气、健脾理湿”的治疗理论,强烈推荐用于治疗UC的方剂。然而,确切的分子药理学机制尚不清楚。
网络药理学被认为是科学揭示疾病、成分和靶点之间关系的有效手段。目前,大多数网络药理学研究通过在TCMSP数据库中筛选口服生物利用度和药物样特性来鉴定草药的生物活性化合物。然而,筛选出的化合物是否真的存在于草药或草药制剂中尚未得到实验验证,更不用说这些化合物在吸收后在靶点组织中的暴露情况。因此,我们通过LC-IT-TOF-MS根据其在UC小鼠结肠中的实际暴露量确定了生物活性化合物,并通过相对含量排名进一步关注SNS的潜在有效成分。最终,KEGG通路分析表明,SNS可能通过调节Th17细胞分化和IL-17信号通路来改善结肠炎。
Th17和Treg细胞分化可以看作是一个标尺的两个相对端。靶向Th17/Treg平衡是治疗UC的一种潜在方法。过量的Th17细胞能够通过分泌IL-17诱导自身免疫和炎症,而Treg细胞介导炎症组织的修复。因此,我们计算了结肠和脾脏中这两种细胞类型的比例。蛋白质印迹、免疫组织化学和流式细胞术的结果为SNS是Th17/Treg平衡的调节因子提供了证据。
我们进一步研究了SNS调节Th17/Treg细胞分化的机制。在通过网络药理学预测获得的五个核心靶点中(图4C),STAT3已被报道调节Th17/Treg分化的平衡。然而,STAT3是网络药理学中常见的预测靶点,我们在PubMed上使用关键字“(STAT3)AND(网络药理学)”进行搜索,在过去三年中产生了554篇相关论文;此外,STAT3在各种免疫细胞中广泛且高度表达(图9A)。总之,STAT3可能不是本研究需要特别关注的靶点。我们将注意力转向了之前的研究,我们发现柴胡皂苷和相应的saikogenins抑制了CYP3A4的催化活性和表达,研究结果于2021年发表在《民族药理学杂志》上。CYP3A4催化胆固醇产生4-羟基胆固醇,这是一种内源性氧化固醇,已被证实会加剧结肠炎。柴胡皂苷是柴胡[伞形科]的重要活性成分,是SNS的君药。这促使我们考虑SNS是否可以通过调节胆固醇代谢来缓解UC。
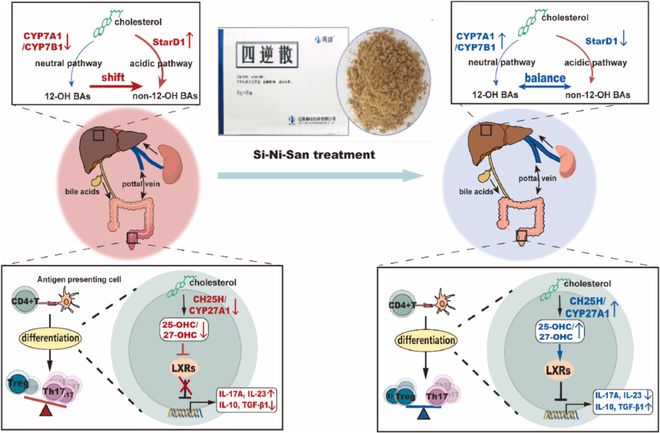
图9 SNS减轻肠道炎症和肝损伤的潜在机制图。作为调节CD4+T细胞分化的转录因子,LXR负向调节炎症Th17分化,促进免疫抑制Treg细胞的分化,从而保护身体免受失调炎症反应引发的损伤。SNS显著促进了结肠中固醇代谢酶的表达,如CH25H和CYP27A1,并显著提高了LXR内源性激动配体氧化固醇(25-OHC,27-OHC)的水平,从而维持了Th17/Treg的体内稳态。此外,SNS可以调节胆汁酸代谢途径关键代谢酶CYP7A1、CYP7B1和StarD1的表达,在炎症状态下逆转BA合成向酸性途径的转变,从而保护肝脏免受结肠炎并发的损伤。
氧化固醇是胆固醇的含氧衍生物。作为LXRs的主要配体,它们可以调节LXRs的活性并影响Th17细胞的分化。LXR(LXRα和LXRβ)是能够影响广泛免疫反应的核受体,可以以T细胞内在或T细胞外在的方式负调控Th17分化。研究还表明,施用合成的LXRs配体可促进Treg细胞在体外和体内的分化。研究表明,在UC小鼠模型和人类IBD中,氧化固醇的分布发生了变化,但研究者尚未阐明氧化固醇在UC发展中的病理作用,很少有研究关注UC发展中失调的氧化固醇LXRs信号传导。因此,在本研究中,我们通过LC-MS/MS测定了氧化固醇含量,并通过检测LXRs和下游基因Abca1和Npc1l1的mRNA表达来研究LXRs的活性。结果显示,与DSS组相比,SNS给药后Lxr、Npc1l1和Abca1的mRNA表达发生了显著变化(图6C)。结肠中的几种氧固醇发生了显著变化,尤其是激活LXR的氧化固醇,如25-OHC和27-OHC,在DSS组中显著降低,而拮抗LXR的氧化固醇4β-OHC的水平在各组之间保持不变。SNS能够逆转这些变化(图7A)。我们进一步在基因和蛋白质水平上测量了氧化固醇合成酶的表达,结果支持了氧化固醇水平的变化(图7B-D)。这些结果表明,炎症性结肠中的氧固醇LXRs信号传导被破坏,SNS通过调节氧化固醇代谢和影响LXRs的表达和活性对UC发挥保护作用。然而,这项研究中仍有一些问题需要回答。我们对氧化固醇LXRs信号传导的研究基于结肠组织,结肠由多种细胞类型组成,如肠神经胶质细胞、肠上皮细胞和免疫细胞。因此,我们还没有探究清楚这种信号的激活是T细胞固有的,还是取决于其他细胞类型。此外,要评估氧化固醇LXRs信号在结肠炎和Th17/Treg分化中的病理生理作用,还有很长的路要走。基因敲除可用于改变氧化固醇代谢或干扰LXRs以供进一步研究。最后,为了阐明SNS的药效物质基础,有必要在体内和体外验证柚皮苷、甘草酸和柴胡皂苷A等前10种化合物在结肠中的疗效。
另一方面,我们观察到UC小鼠的肝脏颜色、质地和胆囊大小发生了显著变化,SNS可以在实验过程中减少这些损伤。我们发现肝脏指标(肝脏指数、AST和AST活性)、结肠炎症指标(IL-6、IL-1β和TNF-α)和DAI之间存在显著的正相关关系(图2)。这些结果表明小鼠结肠疾病活动与肝损伤密切相关。肝脏可能受到肠道菌群失衡和肠道屏障功能受损引起的免疫逃逸的影响。它可以与肠道菌群产生的肠道食物抗原和内毒素接触,并在肠道稳态被破坏后改变免疫状态。在肝损伤期间,免疫细胞被激活以释放大量细胞因子,同时伴有BA的异常分泌。这些肝脏炎症因子和BA信号异常不仅会对肝脏本身造成二次打击,还会损害肠道黏膜屏障功能,影响肠道生态平衡和免疫耐受,形成病理恶性循环,加重UC。
在肝脏中,胆固醇通过中性或酸性合成途径转化为BA,以维持胆固醇稳态。结肠炎破坏了肝脏甾醇稳态,T-CHO和T-BA水平升高,酸合成途径产生的非12-OH BA显著增加,导致结肠炎相关的肝损伤。此外,T-MCA等非12-OH BA的显著增加可通过拮抗法尼醇受体(FXR)导致肠道炎症和上皮屏障损伤,FXR在IBD中抑制炎症并保护肠道屏障。DSS组中性合成途径中关键酶CYP7A1和CYP8B1的基因水平和蛋白质表达显著降低,表明BA的中性合成减少(图8E),StarD1的高表达支持结肠炎小鼠肝脏中BA向酸性合成途径的转化。然而,BA酸性合成途径中的酶CYP27A1的表达在各组之间保持不变。我们推测,DSS小鼠中StarD1的高表达可能会弥补这些现象,从而将结肠炎小鼠的肝脏BA代谢转移到酸合成途径。
总体而言,我们发现SNS可以调节结肠氧化固醇LXR信号传导,抑制CD4+T细胞向Th17细胞的分化,同时促进向Treg细胞的分化(图9)。SNS还通过调节关键酶的表达和逆转BA合成向酸性途径的转变来减轻肝损伤(图9)。因此,我们的新发现表明,SNS可能是治疗肠道炎症和伴随的肝损伤的潜在候选药物。
综上所述,中药SNS通过调节结肠“氧化固醇LXRs”信号传导来维持Th17/Treg平衡,从而在UC中发挥保护作用。它还通过逆转UC小鼠BA合成向酸性途径的转变来改善肝损伤。这些发现深化了我们对SNS在肝脾调节中作用的理解,拓宽了UC的临床治疗选择。
【福利时刻】科研服务(点击查看):、、、、、。咨询加微信:1278317307。




